

פרופ' אלי זמסקי
קיבל את תאריו – בוגר, מוסמך ודוקטור – מהאוניברסיטה העברית. לימד וחקר בה עד שנת 2006, עת פרש לגמלאות כפרופסור מן המניין. המחבר זכה פעמיים בפרס המרצה המצטיין של האוניברסיטה העברית. כיהן כפרופסור אורח באוניברסיטאות הרווארד, יוטה, פנסילבניה, וויסקונסין וצפון קרולינה, שם גם לימד קורסי קיץ. עמד בראש המחלקה לבוטניקה חקלאית בפקולטה לחקלאות של האוניברסיטה העברית. פרסם מאות מאמרים באנגלית ובעברית בספרות המדעית. ערך וכתב עשרים וחמישה ספרים באנגלית ובעברית בנושאים מדעיים, מדע בדיוני וספרי ילדים. כיהן כנשיא מכללת אחוה לחינוך במשך שש שנים והיה המדען הראשי של רשות הטבע והגנים. מכהן כיום כדיקן הפקולטה למדעים מדויקים במכללה האקדמית חמדת בנתיבות.
מערכת השיפה בצמחים היא הספקית האולטימטיבית של המוטמעים לכול אברי הצמח. המוטמעים (הסוכרים, הפחמימות) הם "הכסף העובר לסוחר" בתוך הצמח שבעזרתו הצמח גדל. המטרה של המאמר היא להראות שמנגנון חלוקת המוטמעים בצמחים מקיים כללים של כלכלת שוק חופשי בחלוקת התקציב (המוטמעים) בין הצרכנים השונים.
הובלת מוטמעים בצמח מתרחשת ברקמת השיפה (the phloem) בין מקורות (sources) המייצרים ומייצאים את הסוכרים, לבין מבלעים (sinks) שהם הצרכנים השונים. המקור הראשי שיוצר את המוטמעים ומשווק אותם לכל הצמח הם העלים הבוגרים. עלים צעירים מהווים מבלעים עד אשר הם מתבגרים, מגיעים לפריסה מכסימלית והופכים למקור שמייצא סוכרים. המבלעים השונים הם כול האברים למעט עלי המקור, שורשים, עלים צעירים, קודקודי צמיחה, פרחים פירות, גבעולים. כאשר עלים צעירים המהווים מבלעים הופכים למקור, מתהפך כיוון הזרימה בצינורות הכברה. תופעה זו יכולה לשמש כהוכחה מסיעת לכך שהמנגנון מאפשר שינוי כיוון הזרימה בהתאם למפל הריכוזים בין המקור למבלע (ראו בהמשך). כך גם באברי אגירה, שורשים, או גיאופיטים (בצלים ופקעות) שמשמשים כמבלע חזק של פחמימות בתקופת הגידול והופכים למקור כאשר הצמח פורח (לפעמים עוד בטרם נוצרו עלים חדשים).
ליחסים בין העלים הבוגרים המייצאים מוטמעים, המקור, לבין הרקמות הקולטות את המוטמעים, המבלע, יש חשיבות ראשונה במעלה בקביעת היבול. יחסי מקור/מבלע (source/sink relationships ראו אצל Zamski and Schaffer, 1996), הם בעלי חשיבות עליונה בקביעת מדד היבול (harvest index, HI). מדד היבול הוא מונח חקלאי המחשב את היחס בין משקל היבול לסה"כ משקל הצמח (או משקל היבול לחלקים העל קרקעיים של הצמח כי לעיתים קשה להוציא את כל מערכת השורשים מהקרקע). היבול החקלאי יכול להיות פירות, זרעים, פקעות, עלים וכו'. לדוגמא ה-HI של גרגרי דגנים (תירס, אורז, חיטה), אגוזי אדמה וסלק סוכר נע סביב 0.5 ובתפוח אדמה 0.8 . הטיפוח החקלאי של דגניים, הביא לעליה ב- HI מ- 0.35 ל- 0.5 במאה השנים האחרונות. הנתונים מראים שההשבחה של היבול החקלאי הייתה בכיוון של עלייה ספציפית של היבול ולא עלייה כללית בביומסה של הצמח.
תנועת המוטמעים בשיפה מתנהלת בזרימת מסה (mass flow) ובלחץ לאורך מפל הריכוזים של הסוכרים, כפי שהציע Mϋnch (1930). לכן, ביחד עם המוטמעים נעים בזרימת המסה גם חומצות אמינו, חלבונים, מינרלים, הורמונים ואף גורמים בלתי רצויים כגון וירוסים וחיידקים (Van Bel, 2003). בנוסף לכך, מועברים בזרם המוטמעים גם אותות אזהרה מחלקים שנאכלים על ידי חרקים. השיפה מוליכה גם חומרי ריסוס סיסטמיים כגון קוטלי חרקים וקוטלי עשבים.
מנגנון טעינת הסוכרים לשיפה (sugar loading)
המנגנון שהוצע על ידי Mϋnch בשנת 1930 ונקרא זרימת לחץ (pressure flow, תמונה 2), עדיין נתמך על ידי מרבית החוקרים, למרות שבשנים האחרונות הוכנסו בו מספר שיפורים. המודל מציע שהכוח המניע את זרימת המוטמעים בשיפה הוא מפל הריכוזים בין המקור למבלעים. הסוכרים הנטענים בצנרת, בעלי המקור, גורמים לירידה בפוטנציאל המים ולכן מים חודרים לתוך צינורות הכברה ונוצר בהם לחץ הידרוסטטי המניע את התמיסה כולה בזרימת מסה (mass flow). פריקת הסוכרים במבלעים גורמת ליציאת מים מצינורות הכברה אל העצה. כנימות עלה מנצלות את הלחץ השורר בצינורות הכברה, מחדירות את החדק לתוך הצינורות והתמיסה זורמת לתוך גופן בלחץ (תמונה 3). מכאן אנו למדים גם על חשיבות אספקת המים לעלה, לא רק לשם טרנספירציה, אלא גם לשם יצירת הלחץ של התמיסה בשיפה. המודל מציג את המערכת כסגור הרמטית אבל אחד השיפורים שהוכנסו למודל מציג את המערכת כדולפת/קולטת לאורכה ובאופן זה מספקת מוטמעים לרקמות לאורך המסלול. נמצא שישנה ירידה בלחץ האוסמוטי מהמקור למבלע וכן ירידה בכוח פרוטון מניע וזה מתיישב היטב עם המודל.
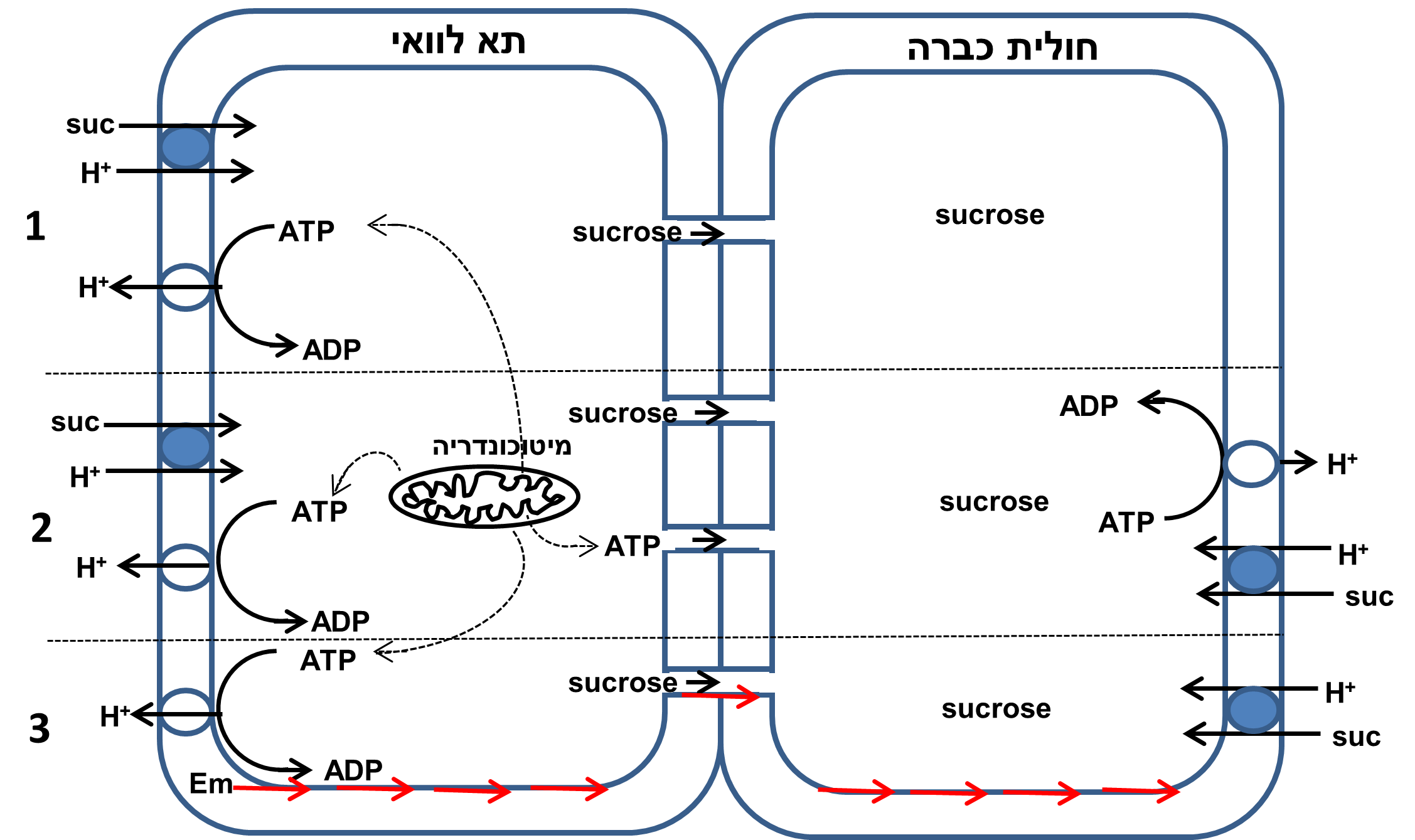
תמונה 1: שלושה מודלים אפשריים לטעינת סוכרוז לתוך צינורות הכברה. 1- סימפורט של סוכרוז ופרוטון כאשר הכוח המניע את כניסת הסוכרוז הוא מפל הפרוטונים (pH חומצי בחוץ). ריכוז הפרוטונים בחוץ נשאר גבוה כתוצאה מפעולת משאבת פרוטון/ATPase הדוחקת פרוטונים החוצה. תמיסת הסוכרים עוברת מתאי הלוואי לחוליות הכברה דרך שפע פלסמודסמטה מסועפים. 2- ATP נוצר על ידי שפע המיטוכונדריה בתאי הלוואי ומשוגר דרך הפלסמודסמטה לחוליות הכברה שם הוא מניע את משאבת הפרוטונים ועל ידי כך מאפשר את פעולת נשא הסימפורט של הסוכרוז והפרוטון הממוקם על ממברנת חוליות הכברה וקולט סוכרוז מהאפופלסט. 3- הפוטנציאל החשמלי (Em) של ממברנת תא הלוואי מועבר דרך הרצף של קרומי התאים (חצים אדומים), דרך קרום הפלסמודסמטה, אל ממברנת חולית הכברה ותורם שם לפוטנציאל החשמלי של הממברנה המניע את הסימפורט של הסוכרוז והפרוטונים פנימה.
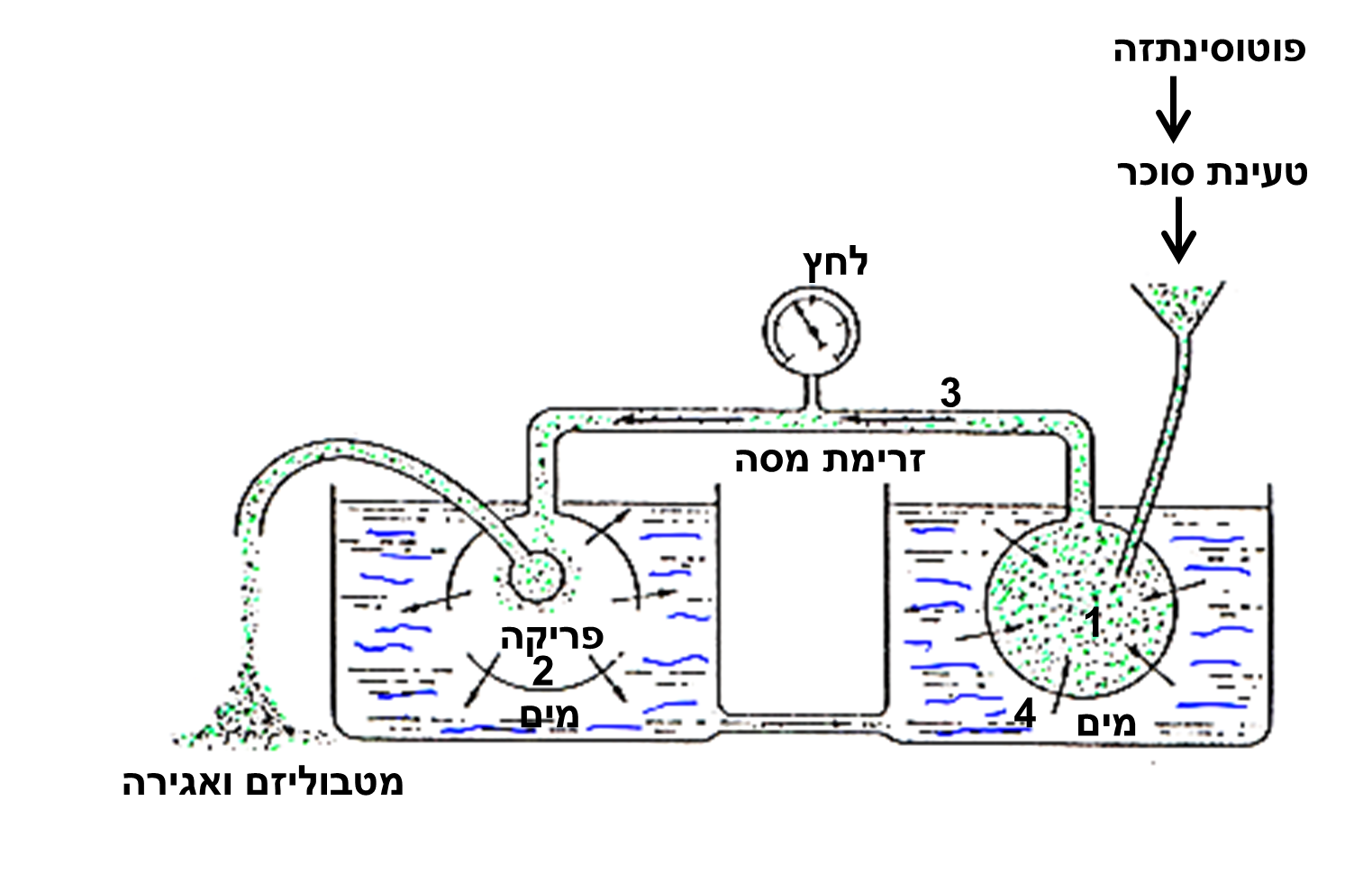
תמונה 2: המודל של Mϋnch (1930) לזרימת לחץ (pressure flow). מערכת השיפה מודגמת בעזרת שני אוסמומטרים הקשורים ביניהם וטבולים באמבט מים. הזרימה היא זרימת מסה של מוהל השיפה בלחץ מאוסמומטר המקור (1) לאוסמומטר המבלע (2) לאורך צינורות הכברה (3). מיכל המים (4) מסמל את העצה המספקת את המים לתוך האוסמומטרים הזרימה ממשיכה כל עוד נמשכת הטעינה של הסוכרים במקור ופריקה שלהם במבלע. הפסקת אחד התנאים האלה תעצור את הזרימה.
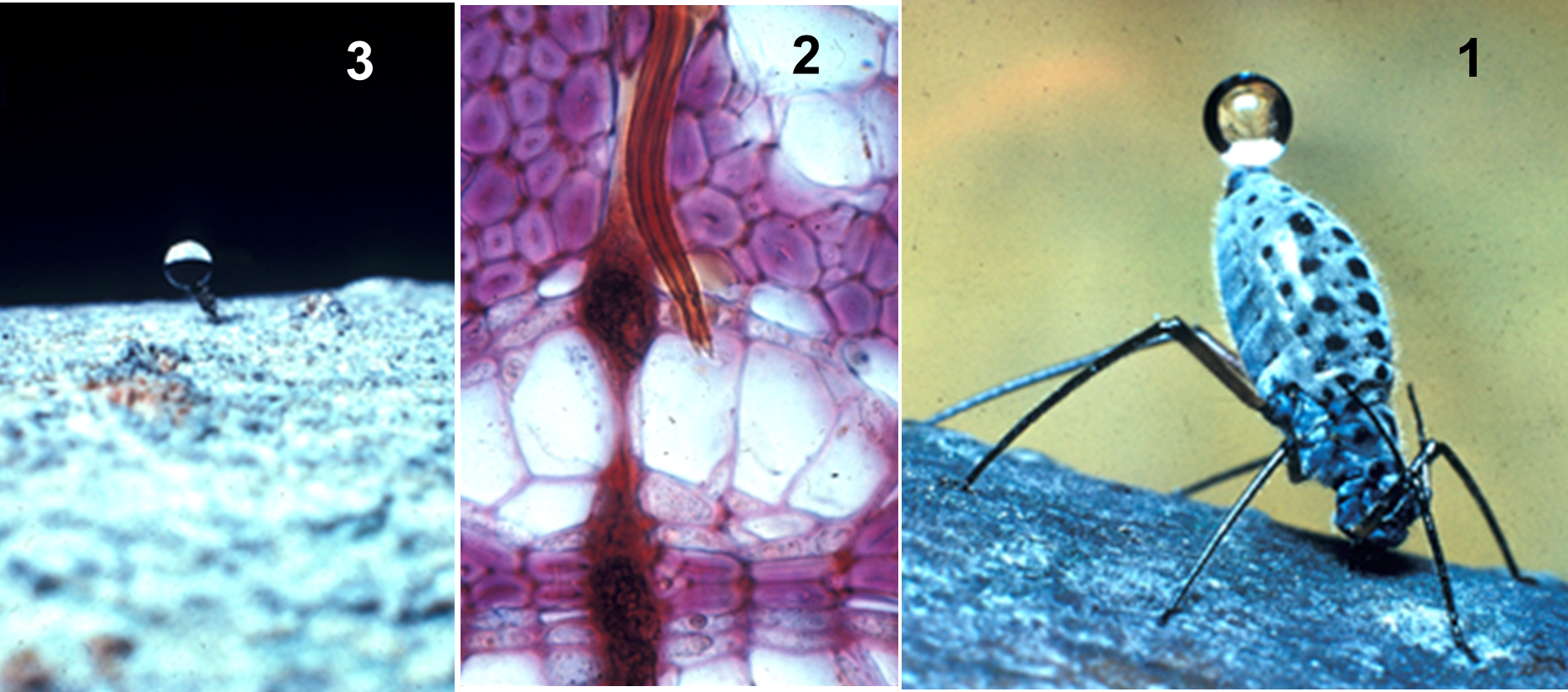
תמונה 3: כנימת עלים ניזונה על ענף של טיליה (Tilia). 1- מבחינים בטיפת "טל דבש" (honeydew) מופרשת מאחוריה של הכנימה. 2- קצה החדק של הכנימה בתוך חולית הכברה. 3- החדק של הכנימה ממשיך להפריש את מוהל השיפה גם לאחר שהכנימה נחתכה והחדק נותר נעוץ בענף. התופעה מוכיחה שמוהל השיפה (phloem sap) מצוי בלחץ וכך הוא גם חודר לגוף הכנימה. התמונות הוענקו למחבר ע" פרופ' מרטין זימרמן (M.H. Zimmermann).
למרות שבמהלך השנים מאז הוצע המנגנון על ידי Mϋnch (1930) ותוצאות ניסויים שבמבט ראשון ביטלו את המנגנון שהוצע, קיימת תמימות דעים רבה לגבי מספר עובדות בהובלת מוטמעים בשיפה-
- מהירות תנועה גבוהה (155-15 ס"מ/שעה. התוכן של כל חוליה מוחלף 5-3 פעמים בשנייה).
- כמות רבה של זרימת חומר יבש (כ- 1.7 גר'/שעה).
- קיים מפל של סוכרים לאורך השיפה מהמקור למבלע (כ- 0.1 מולר/10 מטר).
- קיים לחץ טורגור גבוה (עד 3MPa).
- קיימת תנועה דו-כיוונית בשיפה (אך כנראה לא באותו צינור כברה).
- הובלה קיימת רק בתאים חיים (נדרש קרום התא בחוליות כדי לייצר את הלחץ).
- גורמים המשפיעים על הנשימה משפיעים גם על ההובלה בשיפה (יש צורך ב- ATP).
(phloem unloading) פריקת סוכרים מהשיפה במבלעים
קיימים מספר מסלולים לפריקת הסוכרים מהשיפה בהתאם לסוג המבלע (Zamski, 1996).
- המסלול האפופלסטי (the apoplastic route)- המסלול הזה נפוץ בעיקר לאורך השיפה, בתנועה בין המקור למבלע, בגבעולים ובשורשים. הפריקה/דליפה נעשית אל הדפנות של מערכת ההובלה, צינורת הכברה ותאי הלוואי ומשם נקלטים הסוכרים אל תאי הפרנכימה ונאגרים (במרבית המקרים הופכים לגרגרי עמילן ונאגרים לשימוש חוזר בעת הצורך), או משמשים ישירות ליצירת אנרגיה הנדרשת לתהליכים מטבוליים ולגדילה. המסלול הזה נופץ גם במבלעים שאוגרים סוכרים מסיסים (פרות מתוקים ושורשי אגירה מתוקים כמו סלק סוכר וגזר וגבעולים מתוקים כמו קנה סוכר).
- המסלול הסימפלסטי (the symplastic route)- המסלול הסימפלסטי הוא הנפוץ ביותר בפריקת מוטמעים לתאי מבלע מטבוליים. כך בקדקודי הצמיחה של השורשים והנצר ובגבעולים אוגרי עמילן. ניטרול הסוכרים המסיסים על ידי הפיכתם לגרגרי עמילן, מוריד באופן דרסטי את ריכוז התמיסה ומאפשר המשך פריקה סימפלסטית בכיוון המפל.
- מסלול סימפלסטי שנקטע על ידי צעד אפופלסטי (symplastic pathway interrupted by an apoplastic step)- זהו המקרה הנפוץ ביותר בהתפתחות העובר בזרעים. כידוע העובר נוצר בתוך שק העובר לאחר ההפריה והוא מנותק מרקמות האם וללא קשרי פלסמודסמטה עם הרקמות העוטפות אותו (שריד הנוצלוס, המעטים, עוקץ הביצית). המוטמעים מגיעים בצינורות ההובלה מצמח האם, דרך עוקץ הביצית עד למקום חיבור העוקץ לגוף הביצית בצד החלצה (chalazal region) שם הם נפרקים לאפופלסט. הסוכרים שבאפופלסט נקלטים על ידי תאי מעבר (transfer cells) הממוקמים באינטגומנטים ובשריד הנוצלוס. קליטה מסוג כזה היא קליטה אקטיבית. תאים אלה מפרישים את הסוכרים לתוך שק העובר לטובת העובר המתפתח והאנדוספרם.
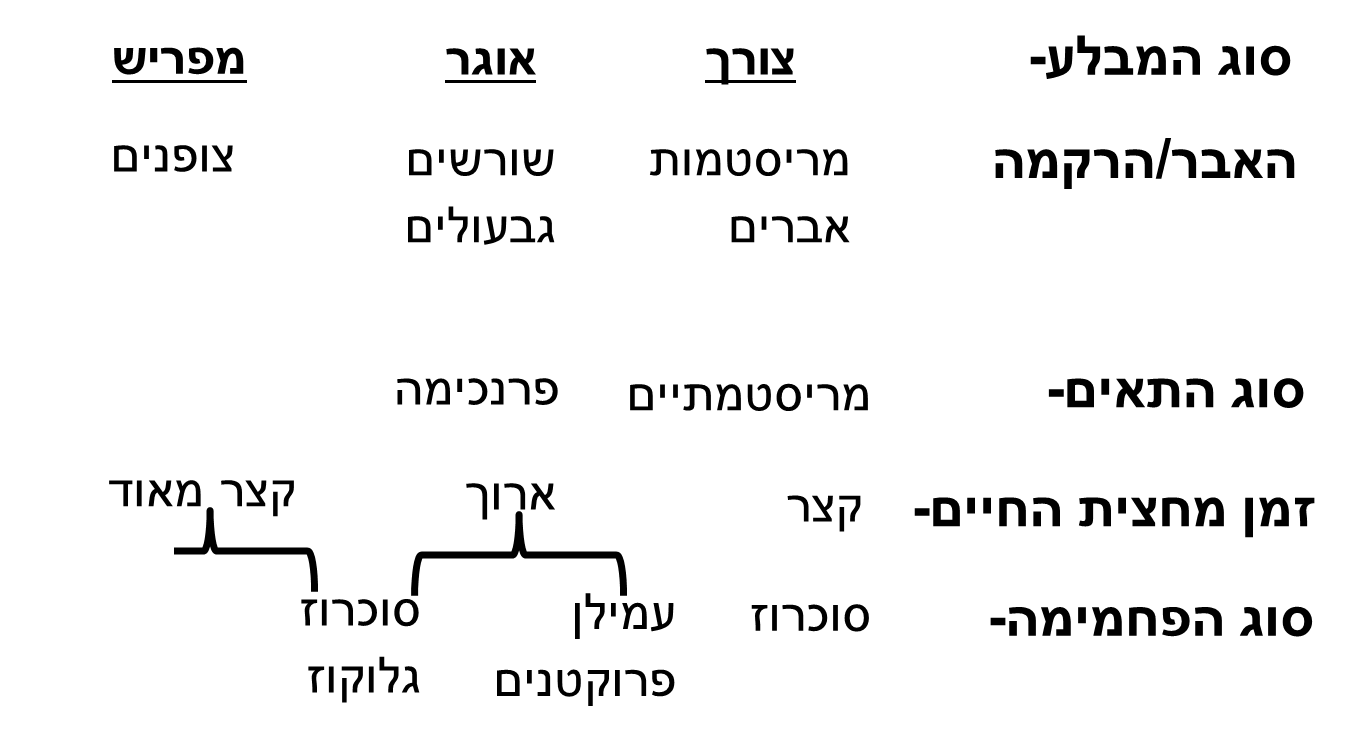
מנגנון הפריקה תלוי בסוג המבלע (תמונה 4) וביכולת שלו לנטרל את הסוכרים המגיעים אליו כדי לשמר את מפל הלחץ. ככול שהמפל בין המקור למבלע הספציפי גדול יותר כך גם תהיה אספקת המוטמעים אל המבלע טובה יותר.
תמונה 4: טבלה המציגה הבדלים בשלושה טיפוסים שונים של מבלעים- צורך לצרכי נשימה וגדילה, אוגר ומפריש וההבדלים העיקריים ביניהם בתפוצה, בסוג התאים, בזמן מחצית החיים של הסוכר בתוך התאים ובסוג הפחמימה העיקרית שבתאים.
במבלעים שאוגרים עמילן (ברקמות אוגרות), שמירת מפל הריכוזים והלחץ תהיה מותנית במעבר הסוכרים לתוך העמילופלסטים ובפעילות האנזימתית של האנזימים המסנתזים שם עמילן. במבלעים מריסטמתיים, תהיה האספקה מותנית בקצב הנשימה וחלוקות התאים. במבלעים שאוגרים סוכרים מסיסים, תהיה האספקה מותנית בקצב קליטת הסוכרים מהאפופלסט, שם הם נפרקים מהשיפה, בקצב קליטתם לתאים ובמידורם בתוך החללית. התהליכים הללו מותנים בפעילות אנזימתית בדופן התא, בציטופלסמה ובחללית. נדרשת פעילות של אינוורטאז חומצי בדופן כדי לפרק את הסוכרוז המגיע מהשיפה, לגלוקוז ופרוקטוז כדי לקלוט אותם פנימה. לאחר מכן התאים יכולים להשתמש בחד-סוכרים לנשימה ויצירת אנרגיה, להחדיר אותם לחללית, או לסנתז מחדש סוכרוז על ידי סוכרוז פוספאט סינתאז (sps) או סוכרוז סינתאז (ss). גלוקוז, פרוקטוז וסוכרוז מוחדרים על ידי נשאים על הטונופלסט לתוך החללית ונצברים שם בריכוזים גבוהים (בסלק סוכר נמדדו ערכים של עד 0.8M). אגירת סוכרים מסיסים בחללית- סוכרוז, או גלוקוז/פרוקטוז, מותנית בנוכחות או אי נוכחות של אינוורטאז חומצי בחללית. אגירת סוכרים מסיסים בחלליות של "המבלע המתוק" נמשכת עד לריכוז מסוים ואז נפסקת כתוצאה מתהליך של וויסות אוסמוטי.
עקרונות לחלוקת המוטמעים בין המבלעים השונים (sugar partitioning and allocation) מתאימים לכלכלה של שוק חופשי
כנאמר קודם לכן, המנגנון המניע את זרימת המוטמעים בשיפה מבוסס על מפל הריכוזים של הסוכרים בין המקור למבלע וזרימת לחץ. אם כך, ככול שהמפל חד יותר הזרימה אל אותו מבלע תהיה נמרצת יותר. על פי אותו עקרון, אם מסיבה כל שהיא יחדל המפל מלהתקיים (לדוגמא הפרי או הפקעת שהתמלאו עד קצה גבול היכולת שלהם לאגור סוכרים), או שהמבלע נושר, תפסק הזרימה לאותו מבלע. המנגנון הזה מבטיח שהמערכת תהיה יעילה מצד אחד, אך גם גמישה כדי לשנות כיוון לעבר מבלע אחר.
צמחים חשים בתנאי הסביבה ומגיבים בהתאם. צמחים מתאימים את חלוקת המוטמעים על פי עקות של גורמים ביוטיים ואביוטיים (ראו סקירה אצל Ruan et al., 2013). כאשר קיים מחסור באספקת מינרלים (בעיקר חנקן וזרחן) לצמח, חל שינוי בביטוי הגנים הקשורים למטבוליזם של הסוכרים בצמח. ישנן הוכחות רבות שהסוכרים משמשים כסיגנלים בביטוי הגנטי של אנזימים הקשורים במטבוליזם של סוכרים. כתוצאה ממחסור במינרלים חלה ירידה בפוטוסינתזה, עלייה בביומסה של השורשים והעלים ועליה בערך הביומסה שורשים/עלים (ראו סקירה אצלHermans et al., 2006 ). המחסור באספקת מינרלים לשורשים גורם לעליה בכמות העמילן בעלים, ירידה בצבירת עמילן בגבעולים לאורך המסלול ועליה באספקה לשורשים. העלייה באספקת מוטמעים לשורשים המצויים בתנאי מחסור במינרלים, נועדה להגביר את צמיחת השורשים כדי לתור אחר המינרלים החסרים.
למרות שמנגנון ההובלה בשיפה נראה על פניו כהגיוני ונתמך על ידי ממצאים רבים, עדיין ישנן סוגיות לא פתורות וקשיים, בעיקר בכל הנוגע לעצים גבוהים (Ryan and Asao, 2014). מסיבות פיזיקליות, ברור שהלחץ בשיפה בעלים של עץ בגובה של עשרות מטרים חייב להיות גבוה יותר מאשר בצמחים עשבוניים כדי להתגבר על ההתנגדויות לאורך המרחק הרב (Thompson and Holbrook, 2003), אבל אין נתונים על כך. כמו כן נמצא שבעצים מסוימים הלחץ בשיפה נמוך מכדי שניתן להסביר את המנגנון (Turgeon, 2010) ושכמעט אין הפרשים בלחץ לאורך המסלול (Lee, 1981). בנוסף לכך, עדיין לא ברור די הצורך מה ההשפעה של עקת מים על מנגנון ההובלה שכן המים נדרשים לשם יצירת הלחץ. Woodruff and Ryan (2014) מצאו כי עקת מים בעצי Douglas Fir (צוגנית מנזיס, Pseudotsuga menziesii חשוף זרע עם תאי כברה בודדים, ללא חוליות כברה!) המתנשאים לגובה של עשרות מטרים גורמת לירידה משמעותית בפוטנציאל המים (13.5% MPa−1), עליה בריכוז הסוכרים המסיסים (0.46 mol l−1 MPa−1) ועליה משמעותית בצמיגות התמיסה (0.27 mPa s MPa−1) בשיפה. מה תהיה ההשפעה של עקת מים על מנגנון ההובלה בשיפה בעצים מכוסי זרע שהם בעלי צינורות כברה?
חלוקת המוטמעים בין העלים, הגבעולים והשורשים היא גמישה כמצופה, ומשתנה בהתאם לתנאי הגידול (ראו סקירה אצל McConnaughay and Coleman, 1999). נבחנה חלוקת המוטמעים בשלושה מיני צמחים חד שנתיים המאופיינים בצימוח מהיר. נמצא שהובלת מוטמעים לשורשים הופחתה ושטח העלים גדל, תחת משטר של עצמת אור נמוכה ואספקת דשנים מוגברת. לעומת זאת לא נמצא שינוי בביומסה של אברי הצמח השונים כתוצאה ממשטרי מים שונים. יתר על כן, אברים אוגרים (גרגרי עמילן, סוכרים מסיסים, או במקרים מיוחדים, חומרי דופן) המהווים מבלעים חזקים בתקופת המילוי והאגירה, הופכים למקורות כאשר הצמח משנה את דפוס הגידול כמו למשל כאשר הצמח עובר מצמיחה וגטטיבית לפריחה, או מתעורר מתרדמה. מכאן נובע שבצמח מסוים מתחוללת תחרות בין המבלעים השונים (מריסטמות, פרנכימה של עצה, שורשים, פרחים, פירות וזרעים) בתקופת הגידול. שורשים ועלים צעירים מהווים את המבלעים העיקריים בתקופה הראשונה של הגידול ואילו פקעות, פרות וזרעים נעשים המבלעים העיקריים בשלב הרפרודוקטיבי (Wardlaw, 1990).
על פי הנאמר קודם לכן ועל בסיס מנגנון ההובלה המקובל בשיפה, ניתן להסיק כי ככול שהמפל בין המקור למבלע הוא גדול יותר, כך גם אספקת המוטמעים לאותו מבלע תהיה טובה יותר. מכאן גם נובע שאם כמות המוטמעים במקור היא נתון קבוע (בכלכלה- "המקורות המוגבלים של התקציב"), הרי שהאספקה נקבעת למעשה על ידי המבלע, או בכלכלה- ע"י הביקוש. ככול שמנגנון הפריקה, בין אם הוא אפופלסטי או סימפלסטי, יהיה יעיל יותר וינקז מתוך צינורות הכברה במבלע את הסוכרים כך גם יישמר המפל בין המקור למבלע. באותו אופן, ככול שינוצלו טוב יותר הסוכרים המגיעים למבלע, אם על ידי מטבוליזם בנשימה, או על ידי מטבוליזם של סינתזת עמילן, או על ידי מידור לתוך החללית, כך גם תמשך הפריקה מהשיפה לאותו מבלע.
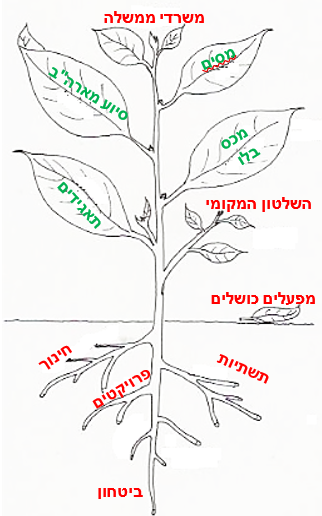
מיחסי מקור/מבלע עולה שהגורם העיקרי המניע את אספקת המוטמעים למבלע הוא "חוזק המבלע" (sink strength). חוזק המבלע נקבע על פי גודלו הפיסי (מספר תאים, מספר פלסטידות וכו') ועל פי הפעילות הביוכימית שלו (פעילות אנזימים). למרות שבמהלך השנים הובעו תמיכה וגם הסתייגות מהשימוש במונח הזה ואף נאמר כי הוא מטעה, השימוש בו מועיל כדי לתאר ביקוש למוטמעים (ראו סקירה בעד ונגד השימוש במונח זה אצל Farrar, 1993). אפשר לתאר אם כן את חלוקת המוטמעים בצמח באנלוגיה למערכת כלכלית שעל פיה מתנהל לדוגמה תקציב המדינה. בתקציב המדינה, כמו בצמח, יש הכנסות (מקורות התקציב, או ייצור סוכרים בפוטוסינתזה) והוצאות (חלוקת התקציב לצרכנים שונים, או מבלעים בצמח). ידוע כי ההוצאה הגדולה בתקציב המדינה היא במשרד הביטחון. באנלוגיה שלנו אפשר לתאר בצמח את מערכת השורשים כצרכן הגדול כאשר הצמח גדל. ההוצאות הגדולות של תקציב הביטחון ("חוזק המבלע") קובעות למעשה שיקבל את הנתח הגדול ביותר בתקציב המדינה (תמונה 5).
המשמעות של מנגנון חלוקת המוטמעים למבלעים השונים, קובעת למעשה שניתן לתאר את החלוקה, באנלוגיה, כמערכת של כלכלת שוק חופשי המושפעת מהיצע וביקוש, כאשר המצרך הוא אחד ובמקרה של הצמח הוא סוכר. המחיר של המוצר נקבע על ידי הביקוש וככול שהביקוש גדול יותר (המפל בין המקור למבלע הוא חריף יותר) כך תגדל האספקה לאותו מבלע (תמונה 6). מבחינה כלכלית אי אפשר להגדיל את ההיצע ללא ביקוש, אבל כאשר גדל הביקוש יגדל במידה מסוימת גם היצור של התוצר (יש בידי האוצר לבקש "תקציב נוסף…"). כאשר נחלשת הזרימה בשיפה (כתוצאה מהחלשות הביקוש), מצטבר סוכר בעלים והפוטוסינתזה תעוכב (Lemoine et al., 2013).

בצמחים מתקיימות תופעות פיזיולוגיות הקשורות ביחסי מקור/מבלע המוכיחות כי חלוקת המוטמעים נעשית על פי הביקוש ולא על פי ההצע:
- עליה בביקוש, למשל התפתחות פרחים ופירות, גורמת לעלייה מסוימת בפעילות הפוטוסינתתית. אך גם זאת באופן מוגבל, כי המקורות הם מוגבלים (דומה לשיקולים הכלכליים הנובעים מ"המקורות המוגבלים של תקציב המדינה", כאשר הוויכוח נסוב בד"כ על השאלה איך מחלקים את התקציב הקיים). צמחים יוצרים בד"כ כמות גדולה של פרחים, יותר גדולה מאשר הם מסוגלים לתמוך בהם עד כדי הבשלת פירות וזרעים. תופעה מוכרת היא שלאחר הפריחה חלה נשירה רבתי של פרחים וגם נשירה של חנטים, בשעה שכמות המוטמעים ("התקציב") אינה מספקת לתחזק אותם. נשירת פירות וחנטים במספרים גדולים מוכרת בעצי פרי, במיוחד בהדרים ובאפרסמון. חשבו ומצאו שעץ בוגר של תפוז מזן ולנסיה יוצר כמיליון פרחים, אבל פחות מאלפית מהם (כ- 800-600) מגיעים להבשלת פירות.
- ירידה בביקוש, למשל ע"י חיגור או הסרת מבלעים, גורמת לירידה בפעילות הפוטוסינתתית.
- מערכת חלוקת המוטמעים בצמח היא גמישה וניתן לציין בתוכה היררכיה של חלוקה על פי עצמת הביקוש. שינויים בהיררכיה מתבצעים כאשר משתנה הביקוש. למשל, בצמח עגבנייה עד שלב הפריחה, המבלע החזק ביותר הוא השורשים, אח"כ עלים צעירים ולבסוף הפרחים. הצמח שומר על ידי כך על כל מה שהוא השקיע בו לזמן ארוך עד שיעשה פירות וזרעים. בתנאי עקה של צמח עגבנייה פורח, נושרים קודם כל הפרחים. יש בכך הגיון רב, כי פרחים הוא יוכל לייצר מאוחר יותר כאשר יוטבו התנאים. לעומת זאת, אם ימותו קודם השורשים, הצמח כולו יבול. ההיררכיה של הצמח משתנה כאשר נוצרים פירות וזרעים. אז בראש הפירמידה של הספקת המוטמעים עומדים הפירות, אח"כ עלים צעירים, אח"כ פרחים ולבסוף השורשים. באופן כזה מבטיח המנגנון את המשך הקיום של המין (תפוצת הזרעים). איך נקבעת ההיררכיה הזו? ע"י מפל הריכוזים! כי זרימת המוטמעים היא טובה יותר ככל שהמפל תלול יותר. כמובן שהצמח איננו "יודע" שכאשר נוצרו הפירות והזרעים "כדאי לו" להסיט אליהם את ההספקה. המנגנון פועל כך שהפירות והזרעים המתמלאים יוצרים ביקוש רב למוטמעים שמסיט אליהם את ההספקה.
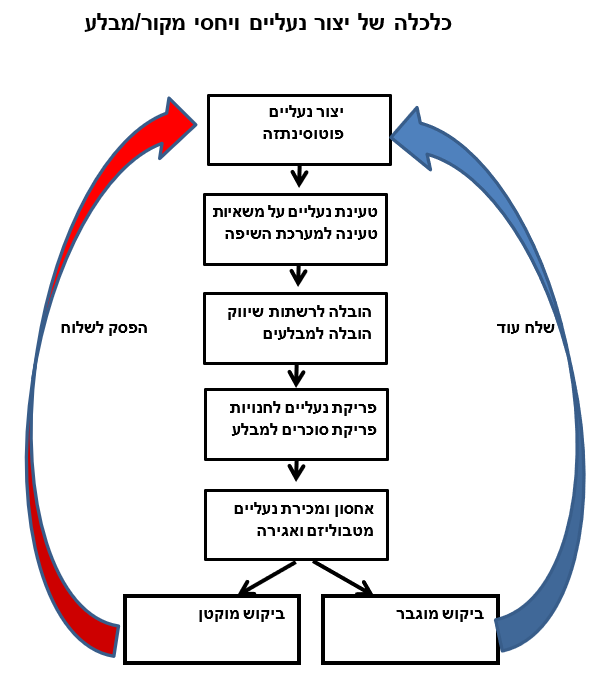
נבחן את הדוגמא הבאה המקשרת בין כלכלה של שוק חופשי ויחסי מקור/מבלע: אם נדמה את הייצור וחלוקת המוטמעים לייצור של נעליים, נקבל התאמות בין כלכלת הנעליים ליחסי מקור/מבלע בצמחים (תמונה 7). נניח שיחדל הביקוש לנעליים בחנויות, יצטברו הנעליים במפעל והוא יחדל לייצר. באופן דומה, אם ברשת חנויות מסוימת יוכרז על מבצע של הורדת מחירים ויגדל שם הביקוש לנעליים, תבוא מיד דרישה מקניין הרשת למפעל הנעליים לשלוח אליו סחורה נוספת… מכאן עולה, שהספקת נעליים לחנויות נמצאת ביחס ישר לרמת הביקוש בכל חנות. באנלוגיה לנעליים, יחסי מקור/מבלע בצמחים מתנהלים על פי עקרונות של היצע וביקוש והמערכת הזו היא גם מספיק גמישה כדי לחדול, להקטין, או להגדיל את ההספקה על פי הביקוש של המבלעים השונים בצמח.
מסקנות חשובות לגבי השבחת היבול
למרות האמור למעלה, שהגורם המבקר את אספקת המוטמעים הוא "חוזק המבלע", בהשבחת יבול הצמחים, יש לקחת בחשבון ששתי המערכות, המקורות והמבלעים של הצמח קשורים זה בזה ולא תיתכן השבחת יבול על ידי טיפול באחד הגורמים בלבד. בנוסף לכך, המשביח חייב לקחת בחשבון מהו התוצר המבוקש. לדוגמה, האסטרטגיה הגנטית תהיה שונה אם המטפח מבקש לפתח זן של תפוחי אדמה גדולים שבו הצמח מניב 5 פקעות בלבד, או 50 פקעות קטנות. בשני המקרים יתכן ומדד היבול, ה- HI יהיה זהה אבל הצמחים יהיו שונים בתכלית והביקוש לתוצרת יהיה שונה. כך גם במקרה של יבול פירות. האם המטרה היא 50 אפרסקים ענקיים לעץ או 200 אפרסקים קטנים.
הצמחים עצמם פתחו במהלך האבולוציה מנגנונים המווסתים את היבול. המטרה האולטימטיבית של כל צמח היא לייצר זרעים שיבטיחו את המשך המין. מן המפורסמות שצמחים שונים מייצרים מספר רב של פרחים. בדרך כלל מספר הפרחים גדול יותר ממה שהצמח מסוגל לייצר מהם פרות וזרעים. לכן, בעזרת בקרה הורמונלית, הצמח "בודק" לאורך תקופת הפריחה ומילוי הפרות והזרעים את יכולותיו "הפיננסיים" כי המקורות הכספיים שלו הם מוגבלים. הבקרה הזו מביאה לידי נשירת פרחים ואחר כך גם נשירת חנטים ונשירת פירות בשלב מוקדם. לדוגמה, עץ בוגר של פרי הדר מזן וולנסיה יוצר כמיליון פרחים אבל בסופו של התהליך מניב רק כ-800-600 פירות. כך מונע הצמח בזבוז משאבים ומבטיח שהפרות שיגיעו להבשלה יכילו זרעים עם עוברים תקינים שיהיו ברי נביטה.
רשימת ספרות
Farrar, J.F. 1993. Sink strength: what is it and how do we measure it? Plant Cell Environ 16:1013–1046.
Hermans, C., J.P. Hamond, P.J. White, and N. Verbuggen. 2006. How do plants respond to nutrient shortage by biomass allocation? Trends in Plant Sci. 11:610–617.
Lee, D.R. 1981. Synchronous pressure-potential changes in the phloem of Fraxinus americana L. Planta 151:304–308.
Lemoine, R., S. La Camera, R. Atanassova, F. Dédaldéchamp, T. Allario, N. Pourtau, J-L. Bonnemain, M. Laloi, P. Coutos-Thévenot, L. Maurousset, M. Faucher, C. Girousse, P. Lemonnier, J. Parrilla and M. Durand. 2013. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 4:272–395.
McConnaughay, K.D.M. and J.S. Coleman, 1999. Biomass allocation in plants: ontogeny or optimality? A test along three resource gradients. Ecology 80:2581–2593.
Münch E. 1930. Die Stoffbewegungen in der Pflanze. Gustav Fischer, Jena, Germany.
Ruan, Y-L., J.W. Patrick, S. Shabala, and T.L. Slewinski. (eds.) 2013. Uptake and Regulation of Resource Allocation for Optimal Plant Performance and Adaptation to Stress. Frontiers in Plant Science 4: article 455.
Ryan, M.G., M.G. and S. Asao. 2014. Phloem transport in trees. Tree Physiol. 34: 1–4.
Thompson, M.V. and N.M. Holbrook. 2003. Application of a single-solute non-steady-state phloem model to the study of long-distance assimilate transport. J. Theor. Biol. 220:419–455.
Turgeon, R. 2010. The puzzle of phloem pressure. Plant Physiol. 154:578–581.
Van Bel, A.J.E. 2003. The phloem, a miracle of ingenuity. Plant, Cell & Environ. 26: 125–149.
Wardlaw , 1990. Tansley Review No. 27. The control of carbon partitioning in plants. New Phytol. 116:341–381.
Woodruff, D.R. and M. Ryan. 2014. The impacts of water stress on phloem transport in Douglas-fir trees. Tree Physiol. 34:5–14.
Zamski, E. 1996. Anatomical and physiological characteristics of sink cells. In: Zamski, E. and Schaffer, A. A. (eds.). Photoassimilate Distribution in Plants and Crops: Source-Sink Relationships. Marcel Dekker Inc. New York. pp. 283–310.
Zamski, E. and A. A. Schaffer, 1996. Photoassimilate Distribution in Plants and Crops: Source-Sink Relationships. Marcel Dekker Inc. New York.